

Половые гормоны
К половым гормонам, в широком смысле слова, относятся, во-первых, гонадотропные, не специфические гормоны передней доли гипофиза, о которых сказано выше и, во-вторых — специфические половые гормоны — фолликулярный, или эстрогенный гормон и гормон желтого тела (прогестерон).
Гонадотропины. По концепции Цондека (1935), в пролане различают два вещества: пролан А — стимулятор созревания фолликулов и пролан Б — стимулятор образования желтых тел. Местом выработки пролана является не только передняя доля гипофиза, но и плацента. Цондек и Ашгейм нашли, что феномен течки у животных под влиянием пролана получается только при наличии функционирующих яичников. Этим окончательно была доказана роль пролана как «мотора» эндокринной функции яичников.
По Цондеку и др., пролан (гипофизарный гонадотропин) имеет большое значение для продолжения беременности. Содержание пролана А в крови на третьем месяце беременности составляет на 1 л 18 000 ME, а пролана Б — 11 000 ME; в последние месяцы беременности — соответственно 19 000 и 8 000. По современной терминологии пролан А называется гонадотропином А или фолликулостимулирующим гормоном, пролан Б — гонадотропином Б или лютеинизирующим гормоном. Гонадотропины вырабатываются не только в гипофизе, но и в плаценте.
Гонадотропный гормон, получаемый из плаценты, сходен с гормоном передней доли гипофиза, но имеет некоторые отличительные свойства, отмеченные И. А. Эскиным (1939, 1941). Так, например, плацентарный гонадотропный гормон (хориальный гонадотропин) отличается от гипофизарного гонадотропина (синоним пролана) тем что, вызывая в яичниках те же изменения, он не способен вызывать их у гипофизэктомированных животных. Секреция плацентарного гонадотропина продолжается в течение всей беременности до момента отделения плаценты.
По новейшим данным, хориальный гонадотропин появляется в моче на следующий день после имплантации яйца, что соответствует сроку не ранее десятого дня после овуляции. В течение первых недель беременности содержание хориального гонадотропина в моче постепенно возрастает и достигает максимума в 150—200 тыс.ME в сутки к шестидесятому дню беременности, считая с первого дня последних менструаций. Выделение больших количеств хориального гонадотропина продолжается около трех недель, после чего его содержание падает до 4—20 тысяч ME в сутки. В дальнейшие месяцы количество хориального гонадотропина снижается еще более: до 200—5000 ME в сутки. К 30-й неделе беременности наблюдается второе повышение продукции гонадотропина в моче, достигающее 40—100 тыс. ME в сутки. Такое повышение может быть кратковременным, и физиологическое значение его неясно. После родов содержание хориального гонадотропина в моче резко снижается, и он полностью исчезает на 5—10-й день после родов. При угрожающих преждевременных родах количество хориального гонадотропина резко уменьшается; при многоплодной беременности и токсикозах первой и второй половины беременности наблюдается повышение содержания хориального гонадотропина в моче (Я. Д. Киршенблат, 1958).
Ф. Бонилла и В. Сальватиерра (Bonilla F., Salvatierra V., 1957) определяли на протяжении всей беременности содержание гонадотропинов в суточной моче реакцией Галли—Майнини. Они нашли, что с началом беременности и до 80-го дня ее выделение гонадотропинов постепенно возрастает; затем оно медленно снижается до 120-го дня, в дальнейшем до конца беременности, оно держится на почти одинаковом уровне, но выше, чем в первые недели беременности. Обычно между 60 и 100-м днем беременности выделение гонадотропинов не падает ниже 10 тыс. ME. При выкидышах уровень гонадотропинов по сравнению с нормой снижается. При содержании гонадотропинов выше 100 тыс. MEпрогноз для сохранения беременности благоприятен.
Гормон задней доли гипофиза (окситоцин, питоцин) выделяется главным образом в родах. При беременности выделение питоцина в кровь, по-видимому, может происходить рефлекторно в результате возбуждения рецепторов матки.
М. Н. Ганусенко (1954) обнаружила, что после расширения шейки матки у беременных (перед выскабливанием слизистой) у некоторых женщин количество питоцина в крови заметно увеличивается. На высоте родовой деятельности питоцина в крови у рожениц больше, чем в начале родовой деятельности.
Эстрогенные гормоны. Фолликулярный, правильнее —эстрогенный гормон продуцируется фолликулами яичника; он содержится в их жидкости. Кроме того, доказано наличие фолликулярного гормона в желтом теле и плаценте. Продуцирование фолликулярного гормона вяичнике женщины носит периодический характер и тесно связано с созреванием фолликулов. В период созревания количество гормона в жидкости и стенках фолликулов достигает максимума, резко снижаясь после менструации.
В желтом теле у женщин фолликулярный гормон содержится в небольшом количестве; максимум его накапливается в период расцвета желтого тела. Одним из доказательств выработки фолликулина плацентой является то, что удаление яичников во время беременности не отражается на кривой выделения фолликулина с мочой.
Наибольшей концентрации в крови фолликулин достигает в фазе пролиферации слизистой матки, особенно к концу ее — перед разрывом фолликула. Этот первый подъем (пик) экскреции эстрогенов наступает, по Е. И. Кватеру (1956), на 6—7-й день после первого дня менструации. В секреторной фазе концентрация фолликулина в крови, по Цондеку, снижается, особенно непосредственно перед менструацией, достигая минимума в первые дни после менструации. Второй подъем (пик) экскреции эстрогенов наступает на 12—15-й день цикла и третий «предменструальный пик» между 20 и 26-м днем цикла. Во время беременности количество фолликулина в крови увеличивается (600 ME на 1 л). Кривая поднимается довольно медленно в первые два месяца, достигая максимума на третьем-пятом месяце, а к концу беременности снова повышается. Максимальное содержание эстрогенного гормона в моче беременных, по Цондеку и Ашгейму, достигает 8—10 тыс. ME на 1 л. Перед началом родов содержание этого гормона в моче резко падает и достигает нормы только через 100—140 часов после родов.
Вначале полагали, что фолликулярный гормон является однородным веществом, которому были присвоены различными авторами названия: фолликулин, эстрин, прогинон. Затем выяснилось, что он состоит из нескольких фракций, которые были названы эстроном и эстриолом. В 1935 г. был выделен еще один дериват — эстрадиол. Более поздние наблюдения Мерриана (Marrian G., 1937) показали, что кривая, составленная Цондеком для тотальных эстрогенов, является правильной, но если учитывать отдельные фракции эстрогенного гормона, то в последние месяцы беременности наблюдается усиленная продукция эстрадиола, а количество свободных эстрона и эстриола снижается, в особенности — эстриола. Полагают, что от половины до ⅔ эстрогенов в крови находится в связанном состоянии.
Обнаруженные в организме женщины три вида эстрогенного гормона — эстрон, эстриол и эстрадиол — не находятся в стабильном состоянии, но способны при различных физиологических и патологических состояниях переходить из одной формы в другую. Метаболизм отдельных эстрионов совершается под влиянием медиаторов, гормонов, а также витаминов и электролитов. В фазе пролиферации менструального цикла в моче находят эстрон, в секреторной фазе, а также во время беременности — эстриол, в фазе десквамации, при месячных — эстрадиол; при аменорее, как правило, эстрадиол не обнаруживается. В последние недели беременности и к началу родов происходит снижение количества малоактивных эстрогенных гормонов (эстрона), количество же биологически высокоактивных (эстриол, эстрадиол) быстро возрастает и держится на высоком уровне во время родового акта. Доказано, что в первую половину беременности (до 5—6-го месяца) в моче обнаруживается эстрон (С. Е. Файермарк, 1946), после этого срока начинается выделение с мочой эстриола; в моче беременных 7—8 месяцев и рожениц, как правило, обнаруживается эстрадиол (Е. И. Кватер и И. И. Каганович, 1947; Рейнольдс С. — Reynolds S., 1949; Е. И. Кватер, 1956).
При преждевременных родах, маточных кровотечениях, отслойке плаценты обнаруживается значительная и длительная экскреция эстрадиола (С. Е. Файермарк). Данные о метаболизме эстрогенных гормонов были использованы для терапии нарушений беременности и некоторых других видов патологии женской половой сферы (Е. И. Кватер, 1956), для чего были предложены способы перевода одних форм эстрогенного гормона в другие.
Колебания уровня гормонов крови в течение беременности зависят отчасти от инактивизации гормонов путем метаболического превращения их в продукты распада, связывания их протеинами и выделения их из организма. В метаболизме стероидных гормонов принимает участие печень (Сегалов — SegaloffА., 1946; Липшюц—Lipschütz, 1944). Инактивизация, или метаболизм, стероидных гормонов происходит также в матке (Смис и Смис; Smith a. Smith, 1938). Именно в матке происходит превращение эстрона в эстриол. Однако роль матки в метаболизме эстрогенов еще окончательно не выяснена.
Содержание эстрогенов при токсикозах первой половины беременности обычно не превышает уровня при нормальной беременности. При токсикозах второй ее половины содержание эстрогенов в крови и моче увеличивается.
При низком содержании половых гормонов возбудимость центральной нервной системы, в частности коры головного мозга, снижается, и раздражения, идущие извне, становятся подкорковыми.
Экспериментальные исследования (Е. А. Какушкина, 1947) показали, что эстрогены изменяют ацетилхолиновый обмен в нервной ткани. Возбудимость ЦНС тесно связана именно с этим (ацетилхолин) медиатором.
Косвенное представление о насыщенности организма эстрогенами дает цитологическое исследование влагалищных мазков. С помощью этого исследования ряд авторов (А. Д. Баландин и соавторы; 1953) показали, что насыщенность организма эстрогенами значительна с ранних сроков беременности; но особенно она нарастает в конце беременности и к началу родов. После родов уровень эстрогенов быстро снижается.
Искусственное введение больших доз эстрогенов в поздние сроки беременности не вызывает прерывания ее по той причине, что при недостаточном количестве прогестерона под влиянием инъекций эстрогенов происходит усиленная продукция прогестерона, что незамедлительно сказывается на усиленном выделении прегнандиола (Смис, Смис и Гурвиц; Smith О., Smith G. a. Hurwitz, 1946). Пероральное введение диэтилстильбэстрола с успехом применялось при угрожающих выкидышах, причем отмечалось увеличение количества прегнандиола (М. Г. Сисьмий, 1952). Такое же действие, но при ранних сроках беременности, оказывает применение тиреоидной терапии.
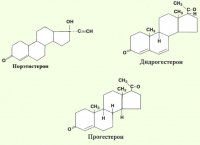
Гормон желтого тела. Гормон желтого тела, прогестерон, вырабатывается в яичниках лютеиновыми клетками желтого тела; небольшое количество его образуется также в лютеинизирующихся клетках зрелых, а частью незрелых фолликулов еще до наступления овуляции. С четвертого месяца беременности большие количества прогестерона продуцируются плацентой. Прогестерон образуется также в коре надпочечников. Прогестерону приписывают две основных функции: 1) влияние на слизистую оболочку матки в смысле перевода фазы пролиферации в секреторную (прегравидную) фазу; 2) протективное влияние на матку и оплодотворенное яйцо, причем он создает условия для имплантации и правильного питания яйца. Прогестерон понижает возбудимость матки и подавляет маточные сокращения, способствует росту матки во время беременности, стимулирует развитие железистой ткани молочной железы.
Тормозящее влияние желтых тел на овуляцию установлено опытами с удалением у морских свинок и коров желтого тела, после чего отмечалось наступление течки и овуляции. Деятельность гормонов желтого тела зависит от лютеинизирующего гормона, вырабатываемого передней долей гипофиза. Гормон желтого тела получил в свое время различные наименования: прогестин, корпорин, L-фактор, лютин и лютеостерон. Теперь его называют гормоном желтого тела — «прогестероном».
По Рейнольдсу (Reynolds S., 1949), выработка прогестерона соответствует возникающим потребностям плодоношения; возможно, что прогестерон образуется еще до момента овуляции. Прогестерон, по-видимому, играет роль в разрастании лютеиновых клеток. Изучение продуктов распада прогестерона показывает, что перед овуляцией у женщин выделяется прогестероноподобная субстанция (Смис, Смис и Гаулд; Smith О., Smith G., Gould А, 1943).
Биологической реакции, специфичной для прогестерона, не существует. Доказывается наличие прогестерона нахождением продуктов его распада, в частности прегнандиола. Прегнандиол выделяется с мочой у небеременных женщин во второй половине менструального цикла в течение нескольких дней по 1—8 мг в сутки и исчезает за 2—3 дня до наступления менструации. Во время беременности прегнандиол выделяется с мочой в больших количествах, но нерегулярно. В ранние месяцы беременности он выделяется циклично наподобие выделения его во второй половине менструального цикла. После 5-го месяца беременности прегнандиол выделяется в больших количествах. Таким образом, продукция больших количеств прегнандиола происходит в те месяцы, когда функции желтого тела почти прекращаются, что и является доказательством продукции прогестерона плацентой.
Во время беременности в течение первых четырех месяцев ежедневно выделяется в среднем от 5 до 10 мг прегнандиола, с пятого месяца беременности, это количество достигает 25 мг в сутки; дальнейшее нарастание происходит медленно и к концу 9-го месяца доходит до 54 мг и даже 120 мг в сутки (Камерон, Веннинг Е.; Т. Cameron, 1948; Venning Е., 1946).
Как говорилось выше, в течение первых четырех месяцев беременности прогестерон продуцируется желтым телом, а позднее плацентой. Количество прегнандиола уменьшается за 2 недели до родов. Через 3-4 дня после родов прегнандиол из мочи исчезает. Выделяемый с мочой продукт распада прогестерона является по своему строению натрий-глюкуронат-прегнандиолом. Роль матки в метаболизме прогестерона выяснена мало. Известно, что превращение прогестерона в прегнандиол может происходить у женщин с удаленной маткой, но для этого необходимо наличие яичников (Джонс и Те-Линде; Jones G. а. Те Linde R., 1941).
Р. Шерман (Sherman R., 1959) определял прегнандиол в моче хроматографическим методом у 51 беременной. Он нашел, что количество прегнандиола постепенно увеличивается с 7 мг при 7-недельной беременности до 43 мг в 40 недель. До 20-й недели количество прегнандиола медленно нарастает, затем, до 30-й недели, быстро увеличивается и с 32-й недели держится на одном уровне. Повторнобеременные до 20 недель выделяют значительно больше прегнандиола, чем первобеременные, в период с 20 до 30 недель — столько же, свыше 30 недель — меньше, чем первобеременные. Установлена определенная зависимость между содержанием прогестерона в плаценте и экскрецией прегнандиола. Выделение прегнандиола зависит от веса плаценты и от веса плода. В некоторых случаях самопроизвольного выкидыша экскреция прегнандиола падает за несколько недель до начала кровотечения.
С. X. Хакимова (1957) у женщин с нормальной беременностью производила определения количественного содержания эстрогенов и прегнандиола в динамике. Установлено, что во второй половине беременности эстриол составляет основной вид эстрогенов в моче, преобладает перед родами и в родах, достигает к концу беременности 2000—3000 гамм, на 7-й день после родов — 34—95 гамм. Содержание эстрадиолатакже повышается к концу беременности (257—412 гамм). Роды сопровождаются снижением количества эстрадиола до 85—170 гамм. На 7-й день после родов количество его в моче значительно возрастает. Количество эстрона в моче (между 17 и 36 неделями беременности) колеблется в пределах 16—111 гамм в сутки. В период «предродового максимума» (С. X. Хакимова) эти цифры возрастают до 231 — 613 гамм, а в родах снижаются (62,4—266 гамм). Со 2—7-го дня после родов он в моче не обнаруживается.
Для прегнандиола характерно раннее появление предродового максимума (на 34 — 35-й неделе). В дальнейшем отмечается некоторое снижение его с последующим повышением в родах. На 7-й день после родов его очень мало.
При резком понижении титра гормонов в родах возможна их патология.
|
Похожие новости
Определение гонадотропной функции гипофизаПередняя доля гипофиза продуцирует гонадотропные гормоны — фолликулостимулирующий гормон (ФСГ), лютеотропный гормон (ЛТГ) и лютеинизирующий гормон (ЛГ), которые оказывают влияние на функцию яичника. Поэтому исследование функции гипофиза имеет большое значение для выяснения патогенеза нарушений менструального цикла.Гонадотропные гормоны выделяют из мочи пОпределение содержания хорионического гонадотропинаХорионический гонадотропин вырабатывается плацентой (хорионом) — ланггансовыми клетками и клетками трофобласта. Хорионический гонадотропин в основном обладает эффектом лютеинизирующего гормона, значительно меньше — фолликулостимулирующего. Многие авторы полагают, что хорионический гонадотропин обладает и лютеотропным эффектом, что обеспечивает функцию желтого тела беременности. Хорионический гонадотропин оказывает дейОпределение содержания прогестеронаУ небеременной женщины гормон желтого тела — прогестерон — продуцируется тека-клетками желтого тела яичников, а во время беременности — синцитиальными клетками хориальных ворсинок плаценты. Имеется связь желтого тела с изменениями во время менструального цикла, причем прогестерон оказывает свое влияние на эндометрий, вызывая разрастание секреторных желез (секреторная фаза цикла) тольОпределение содержания эстрогеновЭстрогены продуцируются яичниками (внутренней оболочкой покрышки), а во время беременности — и хориальными элементами плаценты. Эстрогены вырабатываются также в гранулезо-клеточных опухолях яичников. По первичному строению эстрогены являются производными холестерина. Химическая структура эстрогенов основывается на циклопентанопергидрофенантреновом ядре, общем для всех стероидов.Феномен «зрачка»Феномен «зрачка» основывается на увеличении количества слизи в канале шейки матки в дни, предшествующие овуляции (эстрогенная насыщенность), что при 28-дневном цикле соответствует 12—14-му дню цикла. Динамика изменений в шейке матки в течение нормального менструального цикла была изучена О. П. Голубевой. Установлено, что отверстие матки с 8— 9-го дня цОпределение уровня половых гормонов и их метаболитов в моче и кровиДля определения уровня половых гормонов и их метаболитов в моче и крови применяются следующие лабораторные методы исследования: Определение содержания эстрогенов. Определение содержания эстрогенов в моче. Биологический метод. Определение содержания эстрогенов в моче. Химический метод. Определение содержания прогестерона. Определение содержания хорионического гонадотропина. Определение содержания андрогенныГормон желтого тела и гестагеныГормон желтого тела (прогестерон, или лютеогормон) и вещества, обладающие аналогичным действием, называют гестагенами. Прогестерон, как и эстрогены, относится к стероидам. Прогестерон вырабатывается лютеиновыми клетками желтого тела яичника, которое образуется после овуляции. Во время беременности прогестерон продуцируется плацентой. Может выделяться и корковым веществом надпочечных желез.Эстрогенные гормоныЯичники—половые железы — являются местом образования половых гормонов — эстрогенов (от греч. oestrus—течка) и прогестерона. Эстрогенные гормоны вырабатываются клетками внутренней оболочки покрышки фолликула (tunica interna theca folliculi), прогестерон &mdashГормональная функция яичниковГормональная функция яичников заключается в выделении следующих гормонов: эстрогенов, прогестерона и андрогенов. Эстрогены выделяются клетками внутренней оболочки покрышки фолликула и зернистого слоя. Из натуральных эстрогенов получены эстрадиол, эстрон и эстриол. Эстрадиол — самый активный гормон яичника, эстрон — в 25 раз, а эстриол в 200 разНейро-гуморальная регуляция менструального циклаВ регуляции менструального цикла участвует пять звеньев: кора большого мозга, подбугорная область, гипофиз, яичники, матка. Действие половых гормонов максимально проявляется на матке и яичниках. Функция половой системы регулируется корой большого мозга. Однако локализация центра, регулирующего функцию половой системы, не установлена. Доказано, что удаление коры большого мозга у жи |







Посетители, находящиеся в группе Гости, не могут оставлять комментарии к данной публикации.

Все права защищены
| Информация для специалистов здравоохранения ! | Соглашение об использовании |